
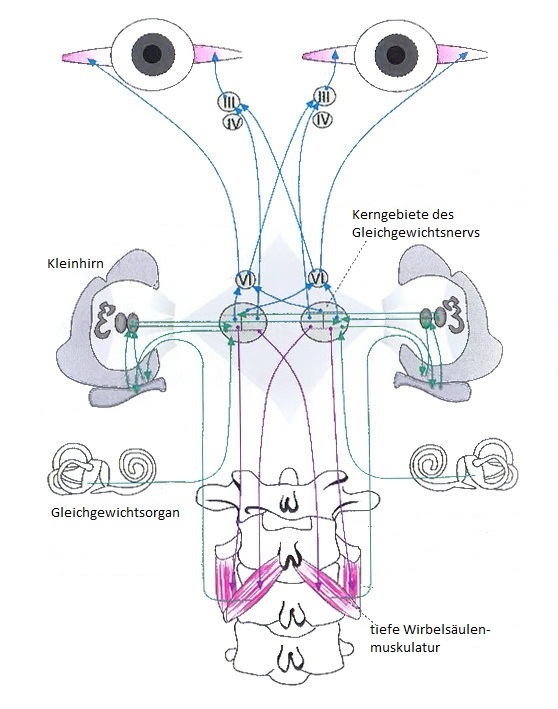
Die motorische Steuerung und Regelung der Augen (Okulomotorik) als auch die motorische Steuerung und Regelung der Wirbelsäule (vertebrale Motorik), speziell der Kopfgelenke, sind funktionell eng miteinander verschaltet. Zusammen mit dem vestibulären System sind diese maßgeblich für die Aufrechterhaltung des posturalen Systems verantwortlich.
Propriozeptive, vestibuläre und visuelle Informationen werden auf Ebene des Hirnstammes integriert. Über den Lemniskus medialis („Körperfühlbahn“, verläuft im Hirnstamm von den Hinterstrangkernen zum Thalamus), den Fasciculus longitudinalis medialis (größte Assoziationsbahn des Hirnstammes mit einem Fasersystem vom Mesencephalon zum thorakalen Rückenmark) und den Tractus vestibulospinalis wird eine Verbindung von den Vestibulariskernen (Ncl. vestibularis, Teil N. VIII) und den externen Augenmuskeln (Ncl. N. oculomotorii, III; Ncl. N. trochlearis, IV und Ncl. N. abducens, VI) einerseits und der Propriozeption aus der autochthonen Wirbelsäulenmuskulatur andererseits hergestellt. So sind die okulomotorischen Mm. rectus lateralis (N. abducens, VI) und die lateralen autochthonen Muskeln (Mm. intertransversarii) homonym innerviert und ebenso die okulomotorischen Mm. rectus medialis und die intrinsischen, autochthonen Mittellinienmuskeln der Wirbelsäule (Mm. rotatores und multifidi). So bilden die lateralen und die medialen Muskelsysteme jeweils ein homologes Innervationsmuster, wenn die Bewegungsrichtung von den Augen getriggert wird. Wir könnten hier von einem okulozervikalen Reflex sprechen (OCR). Die Augenmotorik stellt somit eine Möglichkeit dar, die unwillkürliche innervierte Wirbelsäulenmuskulatur willkürlich über die Augenbewegungen zu beeinflussen. Dies wird z.B. bei Muskelenergietechniken oder vertebralen Justierungen ausgenutzt.
Wird die Bewegung von einer Kopfrotation getriggert, kehren sich die Verhältnisse um: Eine Rotation des Kopfes nach rechts durch Kontraktion der medialen (HWS-)Muskeln (Mm. rotatores) links und der lateralen (HWS-)Muskeln (Mm. intertransversarii) rechts (gekoppelten Mechanik der Halswirbelsäule) ist daher reflektorisch mit einer Kontraktion des M. rectus medialis des rechten Auges und des M. rectus lateralis des linken Auges verbunden [1]. Wir sprechen hier von dem zervikookulären Reflex (COR). Dieser trägt neben dem vestibulookulären Reflex (VOR) dazu bei, dass die Augen einen Fixpunkt bei einer sich verändernder Kopfposition halten können [1]. Klinisch zeigen sich häufig Besserungen von Konvergenzschwächen nach der Beseitigung primärer oder sekundärer Störungen der Kopfgelenke (z.B. als Folge von Malokklusion oder aufsteigenden Störungen des Achsenorgans) [1].
Bei normaler vestibulärer Funktion trägt der zervikookuläre Reflex (COR) nur unwesentlich zur Stabilisierung der Blickbewegungen bei [2; 3; 4]. Das heißt, der Gain (Verhältnis von Augen zu Kopfbewegung) ist gering [5], wird aber bei eingeschränkter Labyrinthfunktion größer. Einen signifikanten Input für die Kopf- und Augenbewegungskoordination liefert der COR bei Patienten mit Labyrinthzerstöruung bis zu einer Frequenz von 0,5 Hz [6; 3].
Willkürliches Mitführen der Augen in die Richtung der Kopfdrehung (M. rectus lateralis rechts und M. rectus medialis links bei Kopfrotation rechts) wird diesen Reflexmechanismus maskieren [1].
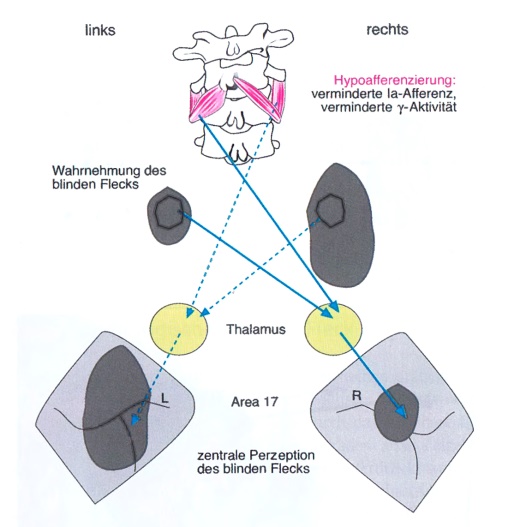
Folgen einer vertebralen Deafferenzierung für das Sichtfeld
50% der visuellen Afferenz eines Auges und alle propriozeptiven Afferenzen kreuzen zur kontralateralen Seite des Thalamus. Bei Hypoafferenzierung kommt es durch Hyperpolarisation (Inhibition) des kontralateralen Thalamus zu einer verminderten kortikalen (perzeptiven) Aktivität. Dies führt als Teilaspekt der thalamischen Funktion (Corpus geniculatum laterale) dazu, dass sich der physiologische blinde Fleck auf der Seite der Hypoafferenzierung vergrößert (Abb. 1) [vgl. 7]. Der physiologische blinde Fleck ist somit ein subjektiv erfassbarer Parameter des Funktionszustands (Summenmembranpotenzial) des Thalamus.
Bei Verbesserung der neurologischen Aktivität wird der blinde Fleck kleiner [7; 8]. Für die Größe des blinden Flecks gibt es keinen verbindlichen Normwert, die Seitendifferenz ist das wichtigste Kriterium. Nach Verbesserung der Afferenzierung, z.B. in Folge einer chiropraktischen Justierung, wird sich der vergrößerte blinde Fleck dem kontralateralen angleichen. Wenn der blinde Fleck links größer ist, müssen die Propriozeption bzw. allgemein die Afferenzen links verbessert werden und umgekehrt.
Wie bereits erwähnt, sind sie intrinsischen, autochthonen Mittellinienmuskeln der Wirbelsäule (Mm. rotatores und multifidi) und die lateralen autochthonen Muskeln (Mm. intertransversarii) homolog zu den medialen bzw. lateralen Augenmuskeln innerviert. Diese Verbindung kommt durch den Fasciculus longitudinalis medialis und den Tractus vestibulospinalis zustande. Eine Rotation der Wirbelsäule, insbesondere des Kopfes nach rechts durch Kontraktion der medialen (HWS-)Muskeln (Mm. rotatores) links und der lateralen (HWS-)Muskeln (Mm. intertransversarii) rechts ist daher reflektorisch mit einer Kontraktion des M. rectus medialis des rechten Auges und des M. rectus lateralis des linken Auges verbunden. Dies entspricht der gekoppelten Mechanik der Halswirbelsäule [1].
Die visuellen Informationen spielen eine entscheidende Rolle bei der Körperorientierung im Raum. Die periphere Retina ist für die dynamische Haltungskontrolle wichtig [10], weil dort das visuelle Raumorientierungssystem seinen Ausgangspunkt hat [11]. Hinzu kommt die Propriozeption der äußeren Augenmuskeln. So konnten Roll, Vedel & Roll [12] 1989 zeigen, dass durch Vibrationsstimulation der äußeren Augenmuskeln mit Frequenzen von 10 bis 80 Hz propriozeptive Afferenzen ausgelöst werden können, die eine Bewegungsillusion der Augen mit dem Eindruck einer visuellen Zielobjektverschiebung mit entsprechenden posturalen Effekten hervorrufen.
Literatur
[1] Garten, H. (2016) Applied Kinesiology – Funktionelle Myodiagnostik in Osteopathie und Chirotherapie. Elsevier, München.
[2] Barlow, D.; Freedman, W. (1980) Cervico-ocular reflex in the normal adult. In: Acta Otolaryngol. 89(5-6), 487-496.
[3] Bronstein, A.M.; Hood, J.D. (1986) The cervico-ocular reflex in normal subjects and patients with absent vestibular function. In: Brain Res. 373(1-2), 399-408.
[4] Sawyer, R.N. et al. (1994) The cervico-ocular reflex of normal human subjects in response to transient and sinusoidal trunk rotations. In: J Vestib Res 4(3), 245-249.
[5] Leigh, R.J.; Zee, D.S. (1999) The Neurology of Eye Movements. In: Oxford University Press, New York.
[6] Kasai, T.; Zee, D.S. (1978) Eye-head coordination in labyrinthine-defective human beings. In: Brain Res 144(1), 123-141.
[7] Carrick, F.R. (1997) Changes in brain function after manipulation of the cervical spine. In: J Manipulative Physiol Ther 20, 529-545.
[8] Carrick, F.R. (1996) Neurophysiological Implicationsin Learning. Onlinepublikation http://www.carrickinstitute.org
[9] Garten, H. (2004) Lehrbuch Applied Kinesiology. Elsevier, München
[10] Berthoz, A. (1974) Oculomotor activity and proprioception. In: Rev Electroencephalogr Neurophysiol Clin 4(4), 569-586.
[11] Levin, L.A.; Arnold, A.C. (2005) Neuro-Ophthalmology. Thieme, New York.
[12] Roll, J.P.; Vedel, J.P.; Roll, R. (1989) Eye, head and skeletal muscle spindle feedback in the elaboration of body references. In: Prog Brain Res. (80), 113-123 & 157-160 (discussion).

Du muss angemeldet sein, um einen Kommentar zu veröffentlichen.